第二节 抗体产生的细胞学基础
一、抗体产生的一般规律
当第一次用适量抗原给动物免疫,需经一定潜伏期才能在血液中出现抗体,含量低,且维持时间短,很快下降,称这种现象为初次免疫应答。若在抗体下降期再次给以相同抗原免疫时,则发现抗体出现的潜伏期较初次应答明显缩短,抗体含量也随之上升,而且维持时间长,称这种现象为现次免疫应答或回忆应答。由于对抗体分子结构研究的进展,发现初次应答产生的抗体主要是IgM分子,对抗原结合力低,为低亲和性抗体。而再次应答则主要为IgG分子,且为高亲和性抗体。TD抗原可引起再次应答,而TI抗原只能引起初应答。对初次和再次应答现象机制的研究,对抗体特异性、多样性、免疫记忆以及对自身抗原而受性机制等问题的研究,都必须以抗体生成的细胞学为基础(图11-1,表11-2)。
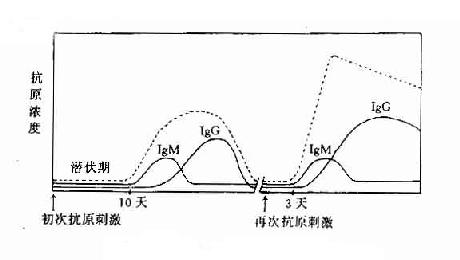
图11-1初次及再次免疫应答
表11-2 初次与再次免疫应答特性
| 特性 | 初次 | 再次 |
| 抗原呈递 | 非B细胞 | B细胞 |
| 抗原浓度 | 高 | 低 |
| 抗体产生 | ||
| 延迟相 | 5~10天 | 2~5天 |
| Ig类别 | 主要为IgM | IgG、IgA等 |
| 亲和力 | 低 | 高 |
| 无关抗体 | 多 | 少 |
二、抗体产生的细胞学基础
抗体产生是由多细胞完成的,Miller等在60年代,首先证明了淋巴细胞是不均一的细胞群。他用早期摘除鸡的胸腺和法氏囊的方法证明了有二类不同的的淋巴细胞,即T和B细胞。前者与细胞免疫有关,后者与抗体形成有关(表11-3)。
表11-3 新生期摘除胸腺及法氏囊对免疫功能的影响(鸡)
| 全身X-线照射 | 周围血淋巴细胞数 | Ig浓度 | 抗体产生 | 移植物排斥反应 |
| 未身X-线照射 | 148 000 | ++ | +++ | ++ |
| 胸腺摘除 | 9 000 | ++ | + | - |
| 法氏囊摘除 | 13 200 | - | - | + |
+阳性反应;-阴性反应
Claman 给经X-线照射小鼠移入同系骨髓细胞(B细胞来源)和胸腺细胞(T细胞来源),然后用羊红细胞免疫,结果证明只有同时移入两种细胞才能产生抗体。因此证明了抗体产生需要T和B细胞共同参予。
Unanue等在70年代又证明了巨噬细胞在抗体形成中的重要作用。他们应用纯化细胞的体外培养技术研究这一问题。根据小鼠细胞对玻璃面或塑料面的粘附性,可将脾细胞分为二种,其一为有粘附性细胞属巨噬细胞(Mφ),另一种为非粘附性细胞属淋巴细胞,包括T和B细胞。当将这二种细胞分别与羊红细胞(抗原)在体外培养时,皆不能产生抗体,只有在二种细胞混合培养时才能产生抗体,自此证明了Mφ也参予抗体的产生(表11-4,5)。
表11-4 T和B细胞在抗体产生中的作用
| X-线照射鼠入的细胞 | 抗体产生 |
| 脾细胞(含有T和B) | ++ |
| 胸腺细胞(T细胞) | ± |
| 骨髓细胞(B细胞) | + |
| 胸腺细胞+骨髓细胞 | +++ |
表11-5 Mφ在抗体产生中的作用
| 体外培养细胞 | 抗体产生 |
| 粘附细胞+羊红细胞 非粘附细胞+羊红细胞 粘附细胞 + +羊红细胞 非粘附细胞 | - - +++ |
表11-6 促进B细胞增殖和分化的细胞因子
| 名称 | 作用 |
| IL-1 IL-4 IL-5 IL-6 IL-2 | 促进B细胞活化与增殖 促进活化B细胞增殖 促进B细胞分化 与IL-1协同 |
在同一时期Gershon等又证明了抑制性T细胞(TS)的存在,对免疫应答起抑制作用。因此TH和TS可视为免疫调节细胞,而TC和TD可视为细胞免疫的效应细胞。通过上述研究,证明了抗体产生需要三种细胞参予,即单核吞噬细胞系、T细胞系和B细胞系。从而否定了过去认为抗体产生是由单一淋巴细胞克隆产生的观点。因此抗体产生不只是涉及抗原与免疫细胞间的相互作用,即对抗原的识别和抗原的激发作用,同时也涉及免疫细胞间的相互作用,,即免疫细胞活化,增殖与分化过程。这二个过程是紧密交织在一起的,为此必须进一步探讨在免疫应答过程中,三种细胞各自发挥什么作用?以及它们之间的相互作用又是怎样进行的。
三、免疫细胞在抗体生成中的作用
上述三类细胞都参予抗体生成过程,但各自发挥的作用不同。现已证明Mφ抗原处理和呈递细胞,无特异识别抗原的功能。T细胞系主要是TH和TS,它们对免疫应答有调节功能,所以是免疫调节细胞,有特异识别抗原的功能。B细胞系既具有呈递抗原的作用又是产生抗体的细胞,也具有特异识别抗原的功能。
(一)Mφ的作用
Mφ在免疫应答的全过程都发挥重要作用,在抗原识别过程中,即在免疫应答的诱导期,它表现为具有摄取、处理加工、存贮和呈递抗原的作用。它活化后还能分泌多种细胞因子,其合成和分泌的IL-1有促进T和B细胞的活化作用。因此,不能认为Mφ只是机械的将抗原决定簇呈递给淋巴细胞,它还具有调节淋巴细胞功能的作用。
抗原性物质进入体内后,必须先经Mφ摄取、加工处理,然后才能呈递给淋巴细胞。Mφ是有吞噬细胞功能的细胞,已证明在其细胞表面有多种受体分子,但无抗原识别受体。它主要是以吞噬、吞饮和被动吸附等方式捕捉抗原,可摄取任何抗原性特质,所以是非特异性的摄取抗原性物质。
摄入的抗原大分子,可在细胞内被降解为许多小肽片段。其中一些免疫原性决定簇可与细胞内的自己MHCⅡ类分子相结合,然后运送至细胞膜表面,形成所谓修饰的自身复合物分子(即异种抗原X+自己MHCⅡ类分子)此即Mφ对抗原的处理和加工过程。
Mφ将这种复合物分子,呈递给有抗原识别功能的淋巴细胞,才能激发免疫应答。不难看出,识别这种复合物的抗原识别受体,必须是既能识别异种抗原X,又能同时识别自已MHC分子。这就是免疫细胞间相互作用的MHC限制性的由来。
(二)淋巴细胞的作用
淋巴细胞具有抗原识别受体,所以T和B细胞都是抗原识别细胞。每一细胞克隆可识别一种抗原决定簇,所以这种识别是有特异性的。B细胞表面抗原识别受体是膜Ig分子,它可识别天然蛋白质抗原分子表面的构像抗原决定簇(即B决定簇),在识别抗原时无MHc 限制性。而T细胞抗原识别受体为异二聚体分子,即TCRαβ,它能同时识别经加工处理的序列决定簇肽片段(即T决定簇)和自己MHC分子,所以有MHC限制性。
四、细胞因子在抗体产生中的作用
细胞因子(cytokines)在抗体产生应答过程中的作用有下述特点:
1.细胞因子的作用既无抗原特异性也无MHC限制性当TH细胞受刺激活化后,它所分泌的细胞因子就可作用于任何抗原特异性的B细胞和任何MHC单倍型(haplotype)的B细胞。
2.在B细胞产生免疫应答的不同时期有不同的细胞因子起作用即B细胞的增殖期与分化期,或Ig的分泌期可有不同的细胞因子在起作用(表11-6)。此外,不同的细胞因子间的组合有的起拮抗作用,有的起协同作用。
3.细胞因子还可作用于旁路(bystander)B细胞使之活化这些B细胞对抗体应答的抗原没有特异性(非抗原特异性B细胞)。它们存在于抗原刺激的特异B细胞周围,在抗原活化特异B细胞时,因产生细胞因子而被活化并产生非特异抗体。
4.在刺激B细胞增殖分化的细胞因子中,有些是来源于巨噬细胞或其他非T细胞所产生(如粒细胞、肥大细胞等)。